Эозинофилы
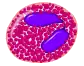
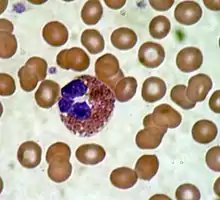
Эозинофи́лы — разновидность лейкоцитов, основная функция которых заключается в борьбе с многоклеточными паразитами. Зрелые эозинофилы имеют ядро, разделённое на две части (двудольное), и эозинофильные гранулы, содержащие белки с цитотоксическими свойствами. Молекулярными маркерами эозинофилов являются белки CD9 и CD35. У здорового человека эозинофилы составляют от 0,5 % до 2 % от общего числа лейкоцитов[1].
| Эозинофил | |
|---|---|
 | |
| Ткань | соединительная |
| История дифференцировки клетки |
Зигота → Бластомер → Эмбриобласт → Эпибласт → Клетка первичной мезодермы → Прегемангиобласт → Гемангиобласт → Гемоцитобласт → Общий миелоидный прародитель → Эозинофильный промиелоцит → Эозинофильный миелоцит → Эозинофильный метамиелоцит → Палочкоядерный эозинофил → Эозинофил |
Название
Названы по свойству окрашиваться эозином (эозинофильность), впервые полученным в Германии в 1873 году Генрихом Каро[2], который дал этому розовому красителю название в честь «розовоперстой Эос»: имя этой богини зари в древнегреческой мифологии было прозвищем его знакомой девушки, Анны Петерс, сестры другого химика[3].
Общая характеристика
.png.webp)
Эозинофилы — относительно немногочисленная группа лейкоцитов, на них приходится 0,5 %—2 % всех лейкоцитов. В крови эозинофилы циркулируют от 30 минут до 18 часов, после чего перемещаются в ткани и пребывают там 10—12 суток. Эозинофилы имеют довольно крупные размеры (от 18 до 20 мкм в диаметре) и двудольное ядро. В цитоплазме имеются крупные (диаметром до 1 мкм) эозинофильные гранулы (так называемые специфические, или вторичные, гранулы). Кроме специфичных гранул зрелые эозинофилы имеют ещё три типа гранул: первичные гранулы, мелкие гранулы, а также липидные тельца[1].
Специфические молекулярные маркеры эозинофилов — белки CD9 и CD35 (рецептор комплемента). Также на поверхности эозинофилов имеются рецепторы для иммуноглобулинов G CD32 и CD16, цитокинов (таких как IL-3, IL-5, GM-CSF) и хемокинов (в особенности эотаксинов). Эозинофилы экспрессируют молекулы главного комплекса гистосовместимости I и II классов, поэтому эозинофилы могут функционировать как антигенпрезентирующие клетки. На поверхности эозинофилов также имеются молекулы адгезии, в особенности, β2-, β1- и β7-интегрины и их рецепторы[1].
Эозинофильность гранул в цитоплазме эозинофилов достигается за счёт главного щелочного белка (англ. major basic protein, MBP). Также в них содержится эозинофильный катионный белок (англ. eosinophilic cationic protein, ECP), эозинофильная пероксидаза (англ. eoxinophilic peroxidase, EPO) и происходящий от эозинофилов нейротоксин (англ. eosinophil-derived neurotoxin, EDN). В гранулах MBP находится в кристаллической форме и образует их сердцевину. ECP, EPO и EDN находятся в матриксе гранул. Специфические гранулы содержат цитокины и ферменты (коллагеназа, эластаза, β-глюкурнонидаза, катепсины, РНКаза, миелопероксидаза). Первичные гранулы заключают в себе кристаллы Шарко-Лейдена, основа которых представлена белком липофосфолипазой. Только у тканевой формы эозинофилов имеются так называемые мелкие гранулы, они содержат в себе ферменты — пероксидазу, кислую фосфатазу, арилсульфатазу и другие. В липидных тельцах содержатся все компоненты, необходимые для биосинтеза эйкозаноидов: арахидоновая кислота и ферменты липоксигеназа и циклооксиненаза. Выделение содержимого гранул происходит путём экзоцитоза[1]. Эозинофилы также секретируют факторы роста — TGFβ, VEGF и PDGF[4][5].
Функции
Главная роль эозинофилов заключается в борьбе с многоклеточными паразитами за счёт внеклеточного цитолиза их клеток. Многие белки, входящие в состав гранул эозинофилов, обладают токсичностью в отношении гельминтов: так, MCP и ECP встраиваются в мембраны их клеток, нарушая их целостность. ECP и EDN являются РНКазами, а потому играют роль в противовирусной защите. Следует отметить, что белки MBP, ECP и EPO токсичны не только для клеток паразитов, но и для клеток самого организма[1]. MBP вызывает дегрануляцию базофилов[6][7] и тучных клеток, MCP может подавлять пролиферацию T-клеток и выделение антител B-клетками, стимулируют фибробласты к выделению слизи и гликозаминогликанов[8].
Когда эозинофил активируется, он начинает выделять разнообразные бактерицидные вещества: активные формы кислорода, перекиси, производные оксида азота NO, цианидов и галогенов. Эти же вещества за счёт окислительного стресса индуцируют гибель клеток посредством апоптоза и некроза[9]. MBP принимает участие в активации тучных клеток и базофилов, поэтому эозинофилы задействованы в развитии аллергических реакций. Кроме того, эозинофилы обладают регулирующей активностью, так как действуют на T-клетки. Эозинофилы задействованы в положительной селекции T-клеток в тимусе, но их роль в этом процессе изучена плохо. Эозинофилы также обладают слабой фагоцитарной активностью. Помимо функций в иммунной системе, эозинофилы регулируют морфогенетические процессы, связанные с половым циклом самок и беременностью[10]. Эозинофилы участвуют в реакции отторжения аллотрансплантата и образовании неоплазий[9].
Эозинофилы, подобно другим иммунным клеткам, выделяют разнообразные цитокины, которые, в частности, участвуют в активации T-хелперов типа Th2. Эозинофилы секретируют широкий спектр цитокинов, в их числе IL-2, IL-3, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12, IL-13, IL-16, IL-18, TNFα, IFNγ, TGFβ, GM-CSF. Помимо того, эозинофилы выделяют некоторые хемокины (эотаксин ССL11, RANTES (ССL5), MIP-1α (ССL3)), эйкозаноиды (лейкотриены, фактор агрегации тромбоцитов (PAF)) и нейропептиды. Эозинофилы чувствительны к хемокинам группы эотаксинов (CCL11, CCL24, CCL26), RANTES и IL-5. С RANTES и эотаксинами эозинофилы взаимодействуют посредством рецепторов CCR1, CCR2 и CCR3. Благодаря эотаксинам эозинофилы могут спонтанно мигрировать к пищеварительный тракт, где они локализуются в lamina propria слизистой оболочки. При менструации и беременности эозинофилы усиленно мигрируют в матку и молочные железы. Некоторое количество эозинофилов перемещается в тимус. В аллергический очаг эозинофилы привлекаются хемокином RANTES, лейкотриенами, PAF и IL-5[11].
Клиническое значение
Состояние, при котором уровень эозинофилов превышает 500 клеток на мкл крови, называется эозинофилией. Эозинофилия чаще всего наблюдается у людей, страдающих от паразитарных заболеваний, некоторых аутоиммунных заболеваний (системная красная волчанка, ревматоидный артрит), а также некоторых онкологических заболеваний, таких как эозинофильная лейкемия, клональная гиперэозинофилия и болезнь Ходжкина. Эозинофилы играют важную роль в развитии астмы, причём количество эозинофилов связано с выраженностью симптомов. Эозинофилы могут вызывать повреждения лёгких у пациентов, страдающих от астмы[12]. Пониженное количество эозинофилов в крови (эозинопения) может быть связано со стрессовыми реакциями, системной красной волчанкой, акромегалией, синдромом гиперкортицизма и приёмом стероидных препаратов[13].
Примечания
- Ярилин, 2010, с. 57.
- Генрих Каро — Биография. www.physchem.chimfak.rsu.ru. Дата обращения: 1 апреля 2020.
- Travis Anthony S. "Ambitious and Glory Hunting . . . Impractical and Fantastic": Heinrich Caro at BASF (англ.) // Technology and Culture. — 1998. — January (vol. 39, no. 1). — P. 105. — ISSN 0040-165X. — doi:10.2307/3107005.
- Kato Y., Fujisawa T., Nishimori H., Katsumata H., Atsuta J., Iguchi K., Kamiya H. Leukotriene D4 induces production of transforming growth factor-beta1 by eosinophils. (англ.) // International Archives Of Allergy And Immunology. — 2005. — Vol. 137 Suppl 1. — P. 17—20. — doi:10.1159/000085427. — PMID 15947480.
- Horiuchi T., Weller P. F. Expression of vascular endothelial growth factor by human eosinophils: upregulation by granulocyte macrophage colony-stimulating factor and interleukin-5. (англ.) // American Journal Of Respiratory Cell And Molecular Biology. — 1997. — July (vol. 17, no. 1). — P. 70—77. — doi:10.1165/ajrcmb.17.1.2796. — PMID 9224211.
- Zheutlin L. M., Ackerman S. J., Gleich G. J., Thomas L. L. Stimulation of basophil and rat mast cell histamine release by eosinophil granule-derived cationic proteins. (англ.) // Journal Of Immunology (Baltimore, Md. : 1950). — 1984. — October (vol. 133, no. 4). — P. 2180—2185. — PMID 6206154.
- Morgan R. K., Costello R. W., Durcan N., Kingham P. J., Gleich G. J., McLean W. G., Walsh M. T. Diverse effects of eosinophil cationic granule proteins on IMR-32 nerve cell signaling and survival. (англ.) // American Journal Of Respiratory Cell And Molecular Biology. — 2005. — August (vol. 33, no. 2). — P. 169—177. — doi:10.1165/rcmb.2005-0056OC. — PMID 15860794.
- Venge P., Byström J., Carlson M., Hâkansson L., Karawacjzyk M., Peterson C., Sevéus L., Trulson A. Eosinophil cationic protein (ECP): molecular and biological properties and the use of ECP as a marker of eosinophil activation in disease. (англ.) // Clinical And Experimental Allergy : Journal Of The British Society For Allergy And Clinical Immunology. — 1999. — September (vol. 29, no. 9). — P. 1172—1186. — doi:10.1046/j.1365-2222.1999.00542.x. — PMID 10469025.
- Rothenberg M. E., Hogan S. P. The eosinophil. (англ.) // Annual Review Of Immunology. — 2006. — Vol. 24. — P. 147—174. — doi:10.1146/annurev.immunol.24.021605.090720. — PMID 16551246.
- Ярилин, 2010, с. 57—58.
- Ярилин, 2010, с. 58.
- Sanderson C. J. Interleukin-5, eosinophils, and disease. (англ.) // Blood. — 1992. — 15 June (vol. 79, no. 12). — P. 3101—3109. — PMID 1596561.
- Zini G. Abnormalities in leukocyte morphology and number (англ.) // Blood and Bone Marrow Pathology. — 2011. — P. 247—261. — ISBN 9780702031472. — doi:10.1016/B978-0-7020-3147-2.00016-X.
Литература
- Ярилин А. А. Иммунология. — М.: ГЭОТАР-Медиа, 2010. — 752 с. — ISBN 978-5-9704-1319-7.